2. THE NUTRITIONAL REQUIREMENTS OF AGARICUS BISPORUS AND COPRINUS COMATUS
2. 1. Choice and identity of the organisms
During the investigations described in this thesis, submerged growth and flavour production of several mushroom species were compared. The choice of the organisms for the more detailed studies was rather arbitrary. Agaricus bisporus and Coprinus comatus were chosen because sufficient amounts of fruit bodies for investigation of the flavour could be obtained easily; the origin of the strains is described in Chapter 6. Unless stated otherwise, our experiments with Agaricus bisporus and Coprinus comatus were done with strains A1 and L, respectively. The identity of Agaricus bisporus, strain A1, was verified by fructification experiments [26]. Coprinus comatus, strain L, did not produce fruit bodies; fructification experiments with this organism have always been difficult. Mounce [109] observed fruit bodies after nine months mycelial growth on a mixture of horse manure and sawdust. The positive results of Lambert (cited by Stoller [135]) were not reproducible. Nevertheless there is no reason to question the identity of our isolate L, especially because the similarity of the odour of this mycelium to that of fruit bodies was confirmed by chemical analyses (Chapters 4 and 5).
Agaricus bisporus, strain A1, needed 20 to 25 days to give the maximum yield in shaken flasks (see Fig. 6). With Coprinus comatus, strain L, the maximum was reached earlier and the yield was three to four times as high as with Agaricus bisporus. Unless stated otherwise, our shaken cultures of Agaricus bisporus were harvested after 21 days, and those of Coprinus comatus after 14 days. Both organisms were thus harvested near the end of the phase of rapid growth.
2.2. Influence of temperature and pH
According to the literature [5, 43, 146] the optimum temperature for mycelial growth of Agaricus bisporus is 23 to 25 °C, while for production of fruit bodies it is 15 to 20 °C. The growth rate of the mycelium quickly decreases above 25 °C. The heat tolerance differs from strain to strain, but most strains hardly grow above 32 °C and die at 34 °C [51]. Nothing is known from the literature about the influence of temperature on the growth of Coprinus comatus.
a After 21 days. bAfter 7 days. c Colony diameter in mm. d Dry weight of mycelium in g/l.
Table 2 shows that for our strains the optimum temperature for the rate of growth on solid as well as in liquid media is 25 °C. The optimum pH for both organisms lies between 6.5 and 7.5 (Fig. 1), which agrees with observations made by Treschov [146] and Volz [50] of other Agaricus bisporus isolates.
In all further experiments the temperature was kept at 25 °C and the initial pH was set at 7. In media with suitable carbon and nitrogen sources the pH decreased during growth. When a carbon source unsuitably for mycelial growth was tested a slight increase of the pH was observed, possibly because the organic nitrogen source was also used slowly as carbon source, resulting in the formation of ammonia. In other experiments no increase of the pH was observed.
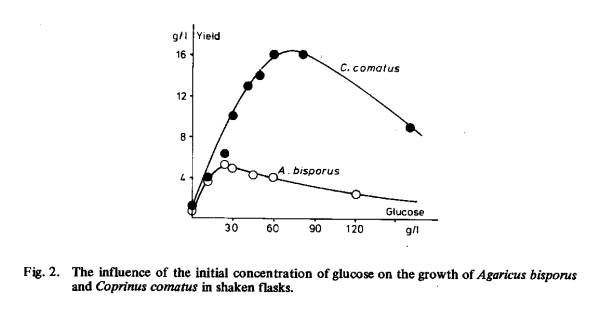
With casein as nitrogen source for submerged growth in shaken flasks, several carbon compounds were tested as carbon and energy sources. The composition of the basal medium is described in Chapter 6.2. When varying the initial concentration of glucose the optimum concentration was found to be between 20 and 30 g/1 for Agaricus bisporus and between 60 and 70 g/1 for Coprinus comatus (Fig. 2).
The yield of Agaricus bisporus could not be improved by gradually adding more glucose during the growth period. This speaks in favour of the view that many other factors than the amount of glucose present limit mycelial growth.
a Concentration: 30 g/1 for Agaricus bisporus and 60 g/l for Coprinus comatus
b Dry weight of mycelium (g/l).
c Compl. = complete medium heat sterilised.
d Sep. = carbohydrate sterilised separately.
e Least significant difference.
+ + = Good growth. + = Moderate growth. - = No growth.
The consumption of other carbohydrates is shown in Table 3. When no carbohydrate was added, growth was not zero as the nitrogen source (casein) could slightly serve as a carbon and energy source. The weight of the inoculum was, at most, 0.006 g/l. Our results are compared in Table 4 with those of investigators who used other strains. As far as they were specified by these investigators, not all the culture conditions were similar. Volz [150], for example, used media with a pH of 4.5 while the other workers used pH values between 6.6 and 7. This again shows that different strains may grow in different conditions. Treschov [146] used static cultures, while the other experiments were carried out in shake flasks. The incubation times varied from 10 to 30 days. The temperatures were between 23 and 30 °C. Treschov [146], Volz [150] and Zarudnaya [164] used synthetic nitrogen sources (asparagine, urea) while Lee [89] used tryptone and we used casein. We may, however, assume that all workers used standardised conditions throughout their experiments, so that their conclusions on the influence of different carbohydrates can be compared. It is clear that many important differences occur between strains of the same species.
Differences in growth occurring when the carbohydrate was sterilised separately or, alternatively, together with the rest of the medium (Table 3) were caused by Maillard compounds; we shall return to this subject in Chapter 2.6.
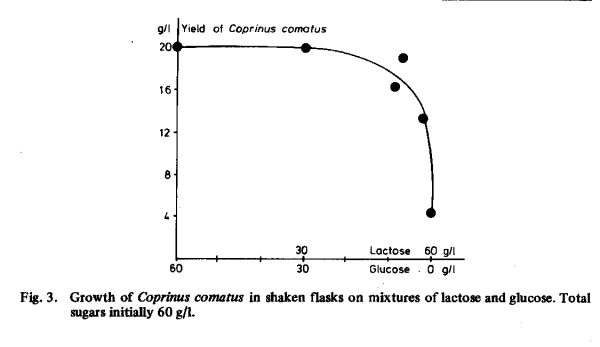
Some of the sugars that were not used by our strains could be made suitable by simple means. Acid hydrolysis of saccharose followed by neutralisation of the acid made it effective for the growth of both organisms (Table 3). The same procedure did not improve the efficacy of lactose. The consumption of lactose by Coprinus comatus was increased by addition of a relatively small amount of glucose (Table 5 and Fig. 3), as had been found in tests with Coprinus lagopus [105]. Galactose was partly consumed in the presence of glucose, but growth on mixtures of glucose and galactose was hardly more abundant than when galactose was omitted.
No galactose was found in the culture medium after lactose had been consumed. If we assume that lactose was hydrolysed in the culture medium, this would imply that galactose could be used completely when it was liberated gradually. Another possibility is that there was no initial splitting of lactose. Glucose might have induced the uptake of lactose from the culture medium. A stimulating effect of glucose on the consumption of saccharose was not observed; nor did glucose appear to have any influence on the consumption of other sugars by Agaricus bisporus.
Metz [168] studied the growth of Penicillium chrysogenum using a carbon source consisting of a mixture of lactose and glucose (25 g and 10 g per litre, respectively). In the initial stage of the fermentation, glucose was used, causing a high growth rate. When the glucose was exhausted, lactose was consumed at a much lower rate, due to the rate-limiting hydrolysis of lactose. The concentrations of glucose and galactose remained very low during this hydrolysis.
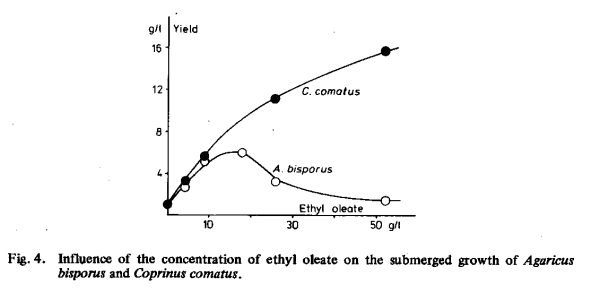
Several non-carbohydrate compounds were tested as carbon and energy sources (Tables 6 and 7). Methanol, ethanol, decane, tetradecane, hexadecane and several salts of organic acids hardly stimulated growth of either organism but often even inhibited it. Sugimori et al. [138] observed good growth of Lentinus edodes on ethanol, but not on salts of organic acids (see Chapter 1). Some fatty acid esters were suitable as carbon and energy sources: methyl oleate, ethyl oleate, methyl palmitate, methyl palmitoleate and, with lower yield, methyl stearate (Table 7). The concentration of ethyl oleate, giving the highest yield observed, was 18 g/1 for Agaricus bisporus and 52 g/1 for Coprinus comatus (Fig. 4).
a Concentration: 18 g/1 for Agaricus bisporus and 36 g/1 for Coprinus comatus.
b Least significant difference.
a All carbon sources 0.06 M.
The effect of higher concentrations of ethyl oleate on the yield of Coprinus comatus was not investigated. From the experimental values listed in Table 8 it may be concluded that oleic acid should be esterified to be suitable as a carbon and energy source, probably because the free fatty acid and even its anions are toxic to the fungi studied.
Apart from being used as the sole source of carbon and energy, the lipids mentioned may play another role in the nutrition of Agaricus bisporus and Coprinus comatus. When added in small amounts to a medium, containing a sugar as the principal carbon source, they often stimulate the growth of Agaricus bisporus, as found by Wardle and Schisler [152]. Such effects were also observed in our shaken cultures of Agaricus bisporus and Coprinus comatus (Table 9). It is interesting that methyl linoleate, which is not suitable as the sole source of carbon and energy, may be used for stimulating the growth of Coprinus comatus in the CME-medium. It is evident that the higher amounts of methyl linoleate added when it was the sole carbon and energy source are toxic for the organism.
Stimulation by lipids varies from strain to strain. Wardle and Schisler [152] reported that some strains of Agaricus bisporus were stimulated by oleates as well as by linoleates, whereas one was not stimulated by lipids at all. Our Agaricus bisporus strain was stimulated by oleic, but not linoleic acid esters.
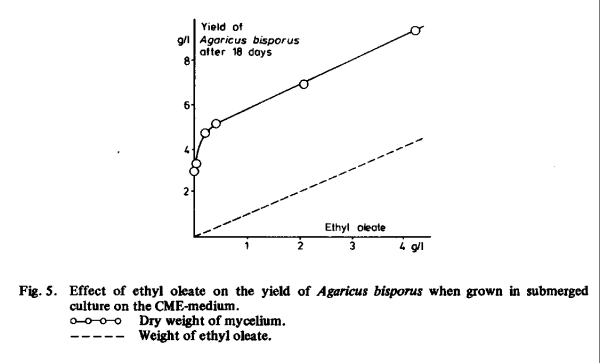
The stimulation by small amounts of ethyl oleate significantly exceeded the weight of the lipid (Fig. 5). Higher amounts (more than 0.5 ml per litre of culture medium) were also taken up, but the increase in yield could be attributed solely to an uptake of lipids from the culture medium. Van den Berg [14] found a curve similar to that in Fig. 5 in tests with Coprinus comatus (strain L).
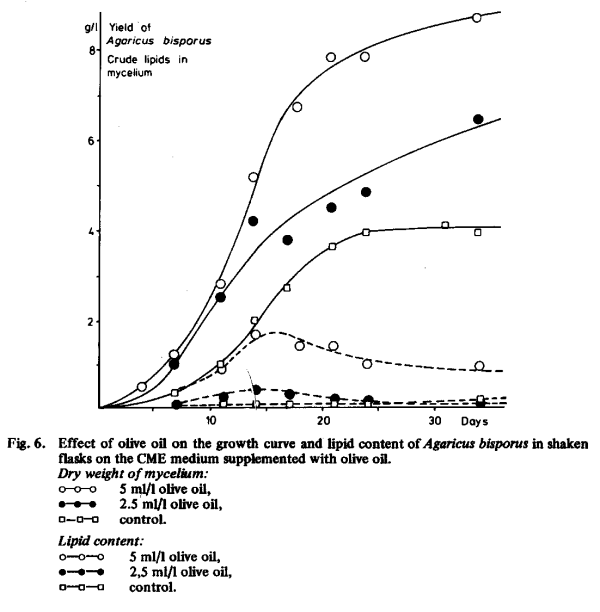
From Fig. 6 it is evident that with olive oil the yield is higher at all stages of growth. The lipid content of the mycelium is maximal after about two weeks, when the yield is approximately half the final yield or higher. Perhaps the lipids are partially incorporated in the cell membrane, thereby facilitating the uptake of nutrients from the culture medium. In later stages of growth the lipids seem to be converted into non-lipid compounds or at least into non-extractable lipid complexes.
The use of lipids in the culture medium changed the composition of the mycelium of Agaricus bisporus. The lipid content of the mycelium increased (Fig. 6) while the nitrogen content decreased, as will be shown in Chapter 2.7.
2.4. Influence of the nitrogen source
With glucose as the carbon and energy source, several nitrogenous compounds were tested as nitrogen sources. Inorganic nitrogen sources, amines, urea and asparagine provided little or no growth (Table 10). Casein was a good nitrogen source, as was a mixture of 17 amino acids having a similar composition as casein. With Coprinus comatus practically the same growth was obtained with the mixture of 17 amino acids as with a mixture of asparagine and phenylalanine; thus this strain does not require any more amino acids. The same conclusion was drawn earlier [26] for Agaricus bisporus (strain A1) from results obtained when its growth on a mixture of asparagine and phenylalanine was compared with growth on a mixture of 20 amino acids, all of which were in the same concentration, while the basal medium contained less glucose (10 g/1 as compared to 30 g/1). From the results presented in Table 10 it is clear, that in more favourable conditions additional amino acids are required for maximal growth of this strain.
a Concentration of nitrogen: 0.06 M (Agaricus bisporus) and 0.12 M (Coprinus comatus).
b Dry weight of rnycelium (g/I).
c 3mM (Agaricus bisporus) and 6 mM (Coprinus comatus).
d Composition, see Table 11.
e Least significant difference.
The amino acid requirements of Agaricus bisporus were investigated by analysing uptake and release from the mixture (Table 11). A medium with the same mixture of amino acids as reported in Table 10 was used. Most of the amino acids were taken up from the medium and there was a particularly high uptake of valine, methionine, leucine, phenylalanine, histidine, lysine and arginine. Probably some of these amino acids cannot be synthesised in sufficient amounts by the strain concerned. This phenomenon was investigated further by using a nitrogen source consisting of mixtures of the same initial concentrations of the amino acids and tryptophane. The initial concentration of tryptophane, which was not determined in the experiments reported in Table 11, was 0.15 g/1. Every amino acid was in turn omitted from the mixture and replaced with an equal amount of leucine. When leucine was omitted, it was replaced with an equal amount of valine. The results are shown in Table 12A.
|
Table 12. Growth of Agaricus bisporus in shaken cultures on mixtures of amino acids in the basal medium. Values not followed by the same letter are significantly different. |
|||
|
|
Amino acids |
Dry weight of mycelium (g/1) |
|
|
A. |
Phe, His, Val, Met, Lys, Leu, Arg, Trp |
2.7 |
DEF |
|
|
------ His, Val, Met, Lys, Leu, Arg, Trp |
0.6 |
A |
|
|
Phe, ------ Val, Met, Lys, Leu, Arg, Trp |
1.7 |
BC |
|
|
Phe, His, ------ Met, Lys, Leu, Arg, Trp |
1.5 |
B |
|
|
Phe, His, Val, ------- Lys, Leu, Arg, Trp |
1.7 |
BC |
|
|
Phe, His, Val, Met, ------- Leu, Arg, Trp |
2.1 |
CD |
|
|
Phe, His, Val, Met, Lys, ------- Arg, Trp |
2.5 |
DE |
|
|
Phe, His, Val, Met, Lys, Leu, -------- Trp |
2.5 |
DE |
|
|
Phe, His, Val, Met, Lys, Leu, Arg, ------ |
3.0 |
EFG |
|
B. |
Asn, |
0.5 |
A |
|
|
Asn, Phe |
2.1 |
CD |
|
|
Asn, Phe, His |
2.5 |
DE |
|
|
Asn, Phe, His, Val |
3.4 |
FGH |
|
|
Asn, Phe, His, Val, Met |
4.3 |
HK |
|
|
Asn, Phe, His, Val, Met, Lys |
4.6 |
K |
|
|
Asn, Phe, His, Val, Met, Lys, Leu |
3.7 |
GHK |
|
|
17 amino acids (see Table 11) |
3.3 |
EFGH |
Omission of phenylalanine, histidine, valine and methionine resulted in a significant decrease in growth, while the need for lysine was doubtful. Leucine, arginine and tryptophane were not necessary. These conclusions were confirmed by the results of another experiment, in which phenylalanine, histidine, valine, methionine, lysine and leucine (concentrations, see Table 11) were added successively to asparagine as the major nitrogen source. The concentrations of asparagine were calculated, so that the total initial concentration of nitrogen was the same within this series of media. The results (Table 12B) showed again the need for phenylalanine, histidine, valine and methionine while the effect of lysine was not significant. Because the effect of phenylalanine was much greater than that of the other amino acids, we may say that the strain concerned had an absolute need for phenylalanine and a relative need for histidine, valine and methionine. Growth with the mixture of asparagine, phenylalanine, histidine, valine, methionine and lysine was significantly better than with the mixture of 17 amino acids.
Frazer and Fujikawa [45] also came to different conclusions about the amino acid requirements of an Agaricus bisporus strain in media with different ratios of glucose and asparagine. In Table 13 the requirements of some strains are compared.
Table 11 also shows uptake and release of amino acids by Coprinus comatus in a medium containing asparagine and phenylalanine. The other amino acids initially present were either impurities in the chemicals or they were introduced with the inoculum. After 24 days the medium contained traces of glycine, alanine, valine, histidine, lysine and arginine in addition to larger amounts of aspartic acid and glutamic acid. The concentration of glutamic acid was so high, that this substance may be an important flavour compound in the culture broth, especially if 5'-GMP has also been formed. The high excretion of glutamic acid was possibly caused by the high concentration of asparagine in the medium, which had shown to result in high yields of mycelium.
+ + = Absolute need for the amino acid.
+ = Growth is promoted by the amino acid.
- = Good growth without the amino acid.
*Essential amino acid for human nutrition.
Our strains of Agaricus bisporus and Coprinus comatus cannot grow with molecular nitrogen as the only nitrogen source (Table 10). The ability of these strains to fix gaseous nitrogen in a complex medium, as found for some mushroom species [53,55,123], was investigated by determination of the total amount of bound nitrogen in a shaken flask containing the CME medium before and after growth; no nitrogen fixation could be demonstrated within the limits of experimental error.
Casein was a better nitrogen source for both organisms than the amino acids, indicating that a complex nitrogen source is better than a defined one, as stated earlier by Styer [136]. This might be caused by slow hydrolysis of casein by extracellular enzymes of the fungi, so that the concentration of free amino acids never becomes as high as in the synthetic mixtures. This theory was supported by the results -obtained in growing the fungi in media with lower initial concentrations of amino acids. More amino acids were added after one or two weeks, so that the amount of nitrogen totally supplied during the growth period was as high as in the experiments reported in Table 10. In this way the growth could be improved significantly; but it was not as high as with casein, probably because the enzymatic hydrolysis of casein supplies the amino acids more gradually than was done in this experiment. It is also possible, that the casein contained one or more unknown compounds stimulating growth.
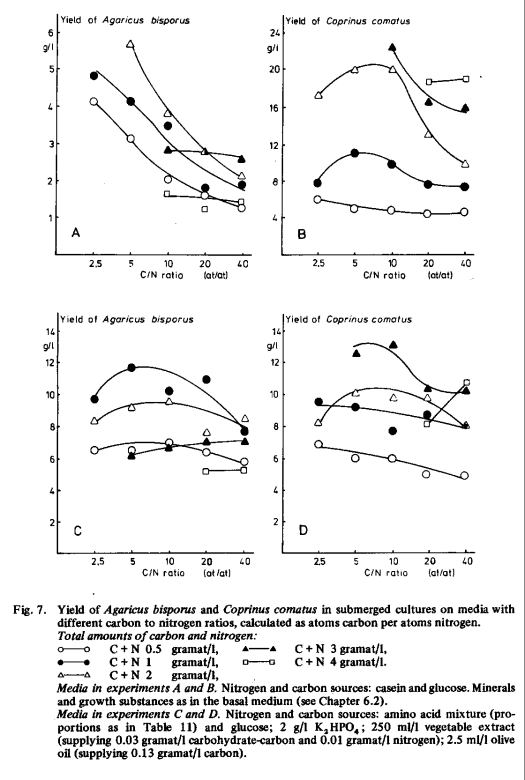
The above results also show that the optimum initial concentration is not the same for all nitrogen sources. This can also be concluded from Fig. 7 where the yield is shown as a function of the carbon to nitrogen ratio with different total amounts of carbon and nitrogen. For the calculation of the C/N ratio the carbon content of casein was ignored because it hardly stimulates mycelial growth (see Table 3). With casein as nitrogen source and glucose as carbon source (Fig. 7A and B) the highest yields of Agaricus bisporus were obtained at low C/N ratios (2.5 to 5); this might be attributed to slow hydrolysis of casein, resulting in low concentrations of available nitrogen. Coprinus comatus grew well with a higher C/N ratio (5 to 10). This suggests that Coprinus comatus hydrolysed casein faster than Agaricus bisporus so that the concentration of available nitrogen became high enough with lower concentrations of casein.
The influence of the C/N ratio was also studied with mixtures of amino acids, thereby providing a readily available nitrogen source. In order to get as high yields as possible, vegetable extract and olive oil were also added to the medium. The results (Fig. 7C and D) show that Agaricus bisporus grew well with C/N ratios between 5 and 20, there being no distinct optimum. When the total amount of carbon and nitrogen was higher than 1 gramat/l the yield decreased. We might attribute this to the sensitivity of Agaricus bisporus to high initial sugar concentrations (Fig. 2), while high concentrations of free amino acids are also less favourable. The C/N ratio resulting in maximal yield of Coprinus comatus growing on the mixture of amino acids (Fig. 7D) was the same as found when the organism grew on casein (Fig. 7B); but the yield was lower. There seemed to be a shift of the optimum C/N ratio to lower values when the total amount of carbon and nitrogen decreased.
Our experiments show, that the optimum C/N ratio may depend on the nature of the nitrogen source and on the total amount of carbon and nitrogen. Thus far we, as most previous workers, have only considered the influence of the C/N ratio on the yield; but the nitrogen content of the mycelium and the uptake of carbon and nitrogen from the medium are also important for the efficiency of the production of mycelium, and these may have their optimum with different C/N ratios. Litchfield et al. [93] found a maximum yield of Morchella hortensis, when the C/N ratio was 10. When this ratio was decreased to 5 a slightly higher yield was observed, but the difference was not significant. With Morchella hybrida Reusser et al. [126] obtained similar results. Shannon and Stevenson [130] demonstrated maximal growth of Calvatia gigantea when the ratio of reducing sugars to nitrogen was 32 to 44 in media based on brewery wastes supplemented with ammonium sulphate. If the non-carbohydrate carbon is ignored, this corresponds to a C/N ratio of 13 to 18, which is higher than in the other observations cited.
2.5. Influence of growth substances
As regards the growth factor requirements of fungi in general we may refer to Fries [47, 48] and Schopfer [129]. Most higher fungi need thiamine as growth factor [47, 77, 80, 151]. In addition, some species need adenine (Fomes officinalis and Poria vaillanti), biotin and riboflavine (Poria vaillanti) or pyridoxine (an unidentified psychrophilic basidiomycete). Only a few species (Morchella conica, Ptychogaster rubescens, Trametes serialis) have been reported to be prototrophic. The growth of Boletus variegatus and Collybia velutipes can be stimulated by fructose- 1, 6-diphosphate [113].
a 400 μg/l.
b 50 mg/l.
c Least significant difference.
Treschov's Agaricus bisporus strain needed thiamine and biotin [46]. All wild-type strains described by Raper et al. [124] only required thiamine and one mutant needed adenine. Ferulic acid promoted growth of another strain of Agaricus bisporus [118]. Coprinus comatus was also shown to need thiamine [33]. In a medium containing yeast extract the strain grew much better, which may be attributed to the need for one or more additional growth substances.
The requirements of our strains for vitamins and nucleobases are illustrated in Table 14. Agaricus bisporus only needed thiamine. The growth of Coprinus comatus was further stimulated by the addition of adenine; the same growth as with thiamine plus adenine was obtained with a mixture of 8 vitamins (including thiamine). Fructose-1,6-diphosphate and ferulic acid did not stimulate the growth of our strains.
In Chapter 2.7 we shall see, that the growth of our strains in various complex media was considerably higher than in the synthetic media studied so far. Possibly there are still unknown factors in complex media which are able to stimulate the mycelial growth.
2.6. Influence of Maillard compounds
If a solution containing a reducing sugar and an amino compound is heated, the development of a brown colour may be observed. This reaction, which was first reported in 1912 by Maillard [99], depends on the pH of the solution: it is rapid at high pH values, still occurs at pH 6 and is inhibited at pH 4. On the other hand the Maillard reaction causes a decrease of the pH of the medium. The chemistry of this reaction is not yet fully understood, although the first steps are well known [57].
During autoclaving of a culture medium, the Maillard reaction may also occur, and the resulting products may effect the growth of micro-organisms. The reaction can be avoided by sterilising the carbohydrate and the nitrogen source separately. If the pH of all media is adjusted to the same value after sterilisation, the effect of Maillard compounds can be studied. Impens and Willam [76] cited several workers who had found a stimulation of the growth of yeasts and lactic acid bacteria by Maillard compounds. Miura et al. [101] found that the lag-phase of Sporobolomyces odorus was shortened by 1-deoxy-l-(L-asparagino)-D-fructose formed during autoclaving of the medium. Impens and Willam [76] also cited some examples of organisms, the growth of which was partly inhibited by Maillard compounds (a Streptococcus species and Bacillus polymyxa).
Impens and Willam [76, 77] investigated the effect of Maillard compounds on the growth of some higher fungi, and found stimulation of Volvariella volvacea and Morchella esculenta, but not of Morchella conica. Growth of some other fungi (Phycomyces blakesleeanus and Aspergillus niger) was also stimulated.
In our investigations the influence of Maillard products was tested in media containing 8 g/1 of casein, 30 or 60 g/1 of glucose and the usual minerals and growth substances (see Chapter 6.2). The pH was adjusted to 7.0 before and after sterilisation. The results obtained with 11 strains are shown in Table 15. Most strains were stimulated by Maillard compounds, although the result was often just below the significant level. The greatest differences were observed in experiments with Armillaria mellea and Lentinus edodes at both sugar concentrations. The yield of Lentinus edodes varied considerably in the absence of Maillard compounds. Five more Lentinus edodes strains were received from the CBS (Baarn, the Netherlands). Their growth was more reproducible; Maillard compounds had a smaller stimulating effect on three of them and none at all on two. A great difference was also found with Coprinus comatus, CBS 150.39, at the lower sugar concentration.
With Agaricus bisporus, strain A1, and Coprinus comatus, strain L, the effect of Maillard compounds was studied with larger numbers of flasks (Table 16). Variations resulting from differences in the shakers on which the flasks were incubated had to be eliminated. The yields and standard deviations of all flasks (maximally 36) incubated on a shaker were expressed as percentages of the average yield in the flasks containing Maillard compounds, on that shaker. The results from all shakers, calculated in this way, were combined. The stimulating effect of Maillard compounds on the growth of Agaricus bisporus, strain A1, was demonstrated conclusively (significance 99%). The growth of Coprinus comatus, strain L, was not stimulated by Maillard compounds, this in contrast to strain CBS 150.39 at the lower glucose concentration (Table 15).
The effect of Maillard compounds was also observed with other carbohydrates (Table 3).
a Compl. = complete medium heat sterilised. Sep. = carbohydrate sterilised separately.
b Significantly lower yield without Maillard products.
c Varying yield.
a Compl. = complete medium heat sterilised. Sep. = carbohydrate sterilised separately.
b Arbitrarily set to 100 %.
Several explanations of the effect of Maillard compounds on t e growth of micro-organisms have been suggested. These include activation of the sugar [116] and activation of thiamine [77]. The inhibition of the formation of a toxic product is also possible. For example, when cystine is autoclaved, decomposition may lead to the formation of substances that are toxic for Lactobacillus bifidus; this decomposition is inhibited when cystine is autoclaved together with glucose [20].
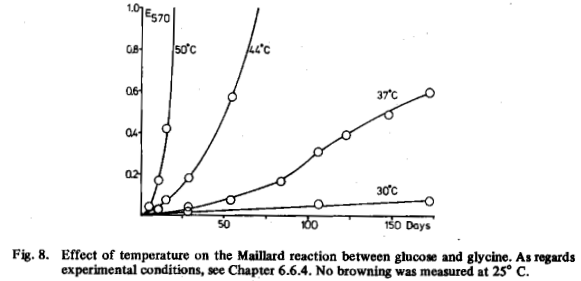
Although the Maillard reaction takes place most quickly at higher temperatures, (60 to 120 °C), it also occurs after prolonged incubation (some months) at 37 °C and it does not stop at 30 °C (Fig. 8). We did not measure any browning at 25 °C, although a pale yellowish colour was visible after 170 days. Thus Maillard compounds may also be found in natural conditions. It does not seem unlikely that many micro-organisms are adapted to grow in the presence of Maillard compounds.
2.7. The use of complex media
Complex sources of nutrients (preferably waste materials) are most often used in large scale cultivation of fungi, since they are cheaper than synthetic media or have to be disposed. In addition, growth of many strains is more abundant in complex media, either because unknown growth substances are present, or because high molecular carbon or nitrogen sources are broken down gradually, so that the concentration of sugars and amino acids does not become excessive. The following waste materials have been used as carbon and energy sources for higher fungi: citrus press water [16], sulphite waste liquor [21, 63, 83, 126], corn steep liquor [63, 125], vinasse [38], molasses [63, 126], soy bean whey [39], soy been oil [125], spent wash, a by-product of the alcohol industries [63], and brewery wastes [130]. As complex sources of nitrogen were used: yeast extract or yeast autolysate [33, 138], corn steep liquor [125], vegetable wastes like cauliflower leaves [79] and brewery wastes [130]. Several higher fungi also utilised simple ammonium salts.
The complex carbon and nitrogen sources mentioned also provided trace elements and growth substances. A small amount of corn steep liquor or yeast extract can also be added as source of growth substances to media with simple carbon and nitrogen sources [52, 59, 60, 93]. Coconut milk was used as a source of growth substances for Volvariella volvacea [6].
a 250ml/l (Agaricus bisporus) and 500 ml/l (Coprinus comatus). b 2g/1. c 4 g/l. d 1 g/l.
e 5 g/l (Agaricus bisporus) and 10 g/l (Coprinus comatus). f 60 g/1.
g 30 g/l (Agaricus bisporus) and 60 g/l (Coprinus comatus). k 20 ml/l. l 20g/1. m 200 ml/l. n 0.4 g N/1.
p 3.9g solids/1. q least significant difference.
For our Agaricus bisporus and Coprinus comatus strains it was necessary to find an organic source of nitrogen (amino acids) and growth substances, all of which were needed by each strain. In Table 17 the growth in several natural media is listed. Malt extract contains carbohydrates, vitamins and a small amount of nitrogenous substance; supplementation with a suitable nitrogen source increased the growth. As might be expected from the results of former experiments, casein was the best supplementary nitrogen source; for Coprinus comatus asparagine was also effective. Surprisingly, also urea stimulated the growth of Coprinus comatus in malt extract. Coprinus comatus also used skim milk after the addition of glucose. Better growth was obtained after addition of technical maltose, a rather impure preparation. The same yield was obtained when glucose and meat or vegetable extract were added to skim milk. Agaricus bisporus cannot exhaust skim milk media completely because it can make a limited use of lactose only, even in the presence of glucose. However, fairly good growth was obtained, especially when technical maltose or vegetable extract were added.
The best growth of Agaricus bisporus was observed when vegetable oil was the carbon source and casein the nitrogen source. Addition of meat extract or vegetable extract stimulated the growth. Coprinus comatus also grew well in these lipid containing media, but not as well as in the skim milk media. Corn steep liquor was not suitable for growth of the fungi studied. Profio is a cheap cotton-seed flour containing a high amount of protein, several minerals, vitamins, some lipids and a limited amount of carbohydrates. When supplemented with more glucose, it was suitable for growth of Coprinus comatus. However, Agaricus bisporus did not grow with Proflo. Single cell protein, prepared from cells of the yeast Pichia pastoris grown on methanol, also promoted the growth of Coprinus comatus while growth of Agaricus bisporus was not abundant with this nitrogen source.
Table 18 shows the conversion of carbon and nitrogen by the mycelium when grown in the media affording the highest yields mentioned in Table 17. Coprinus comatus, grown in medium 9, contained 50% carbon and 8.3% nitrogen. Agaricus bisporus, grown in medium 13, contained 61% carbon and 2.6% nitrogen. This low nitrogen content is related to the high lipid content of the mycelium, grown in a lipid containing medium, as was already mentioned (Chapter 2.3). When grown in a medium based on skim milk (no 7 in Table 17), Agaricus bisporus mycelium contained 10.8% nitrogen. The fungi did not exhaust the media. On varying the concentrations of components of several media listed in Table 17 higher yields were sometimes observed, but the gain in yield was much less than the additional nutrients supplied. Probably these more concentrated media are utilised less efficiently. For an economically feasible conversion of the carbon and nitrogen sources more dilute media should be used.
a Numbers refer to Table 17.
b Expressed in g carbon/l.
2.8. Discussion
The nutritional requirements of our strains of Agaricus bisporus and Coprinus comatus are rather complex, but it may be possible to find less expensive sources of suitable nutrients. The results with Coprinus comatus growing on Proflo plus glucose and on malt extract plus urea, and the ability of Agaricus bisporus and Coprinus comatus to use a vegetable extract (from cauliflower leaves) as a source of growth substances suggest possibilities. Moreover, many other complex media may be investigated.
The examples that were given on the utilisation of sugars, the need for growth substances and the stimulation by lipids show that much variation exists between different strains of the same mushroom species. So far, little or no variation has been found in the optimum pH and temperature.
REFERENCES TO CHAPTER 2
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}