5. PRODUCTION OF FLAVOUR COMPOUNDS BY MUSHROOM MYCELIUM IN SUBMERGED CULTURE
5.1. Brief literature review
The flavour of mushroom mycelium grown in submerged culture was previously studied mainly by sensory observations, mostly based on a limited number of judgements. Sugihara and Humfeld [137] found that Lepiota rhacodes developed a mushroom flavour in submerged culture. The flavour of Morchella mycelium resembled closely the flavour of the fresh fruit bodies [52,79,88,93]. Hamid et al. [63] observed that submerged mycelium of Tricholoma nudum had a pleasant flavour. Torev [144] also reported favourably on the flavour of his cultures. Sugimori et al. [138] found that submerged mycelium of Lentinus edodes had a better mushroom flavour when grown on ethanol than when grown on glucose. Mycelium of Boletus edulis, when crushed or disintegrated, clearly had the smell and taste of the fruit bodies [37].
On the other hand Block et al. [16] did not observe any mushroom flavour in submerged mycelium of Agaricus blazei. Eddy [33] reported that mycelium of Coprinus comatus only had a very weak flavour and Ginterová [54] found that Pleurotus ostreatus produced no flavour at all in submerged culture. She attributed this to the fact that submerged mycelium became monokaryotic, while mycelium in static cultures was dikaryotic. Musilek et al. [167] observed that moist mycelium of Oudemansiella mucida had an agreeable ester odour quite different from the typical mushroom odour of the corresponding fruit bodies.
Only LeDuy et al. [88] used gas chromatographic and spectrophotometric methods to compare the flavour compounds of submerged Morchella mycelium with that of Agaricus bisporus fruit bodies. They detected several differences in the spectra, but did not identify any compound.
5.2. Production of some flavour compounds by higher fungi in submerged culture
After it had been shown that 1-octen-3-ol, 5'-GMP and glutamic acid were important flavour compounds of most mushrooms (Chapter 4), these compounds could be used as test substances in studying the development of a mushroom flavour in submerged culture. Several higher fungi were examined. Table 31 shows the results. The fungi were grown in the CME medium, which contained casein. Because glutamic acid is liberated during hydrolysis of casein, the biosynthesis of this acid by the mycelium was not studied in such cultures.
In addition, the strains of Beauveria tenella were investigated, although Beauveria is not a mushroom genus, because they are the fast growing strains isolated as Agaricus campestris by Humfeld [73] which were later identified as Beauveria tenella by Molitoris [103]. No 1-octen-3-ol was produced by Beauveria tenella, whereas it was detected in the mycelium and in the medium of both Agaricus bisporus strains. This is new support of Molitoris's finding; and it explains the lack of mushroom flavour in the mycelium of Humfeld's strains. In fact, the mycelium had a nut-like flavour, which was not unpleasant, but not at all like that of mushrooms.
The two isolates of Agaricus bisporus did not produce equal amounts of 1-octen-3-ol. Isolate I8 differed from isolate A1 in the appearance of the colonies on solid media, where isolate I8 formed more fluffy colonies than isolate A1. Although isolate I8 gave higher yields of mycelium, the total production of 1-octen-3-ol was considerably lower than that of isolate Al.
Agaricus bitorquis did not produce 1-octen-3-ol in submerged culture, although fruit bodies of this species contain high amounts of this substance. Since the loss of flavour production might have resulted from the strain's having been maintained in pure culture for some years, a fresh isolate from fruit bodies of the same habitat was made. This isolate showed, however, the same behaviour.
Calvatia gigantea, fruit bodies of which contain very large amounts of 1-octen-3-ol, also produced it in submerged culture, where it was released to a large extent into the medium.
The two isolates of Coprinus comatus showed different behaviour. Our own isolate (strain L) produced 1-octen-3-ol, although not very much, while the CBS-strain (150.39) did not produce it at all. Probably Eddy [33] worked with the same strain. Our results agree with his finding that the flavour of the submerged grown mycelium of this strain is very weak.
The two Morchella species released 1-octen-3-ol into the medium but the mycelium contained the substance too. This is consistent with the fact that mycelium of Morchella species grown in submerged culture has almost always been reported to have a good mushroom flavour.
Little or no 1-octen-3-ol was found in submerged cultures of Armillaria mellea, Lentinus edodes, Pleurotus ostreatus and Volvariella volvacea. Of these four species only fruit bodies of Pleurotus ostreatus are known to contain rather much 1-octen-3-ol, while dried fruit bodies of Lentinus edodes contain a small amount (Table 30). No data are available on the 1-octen-3-ol content of fruit bodies of Armillaria mellea and Volvariella volvacea, so that no comparison can be made with the submerged mycelial cultures. The poor production of 1-octen-3-ol by our strain of Pleurotus ostreatus is consistent with the finding of Ginterová [54] that other Pleurotus ostreatus strains produced no flavour in submerged culture.
We have now seen that the absence of a mushroom flavour reported in the literature coincides with the absence of 1-octen-3-ol (Beauveria tenella; Coprinus comatus, CBS 150.39; Pleurotus ostreatus) while species, reported to develop a good mushroom flavour, produced 1-octen-3-ol (Morchella species). Further, the findings of Abbott and San Antonio [1] that fruit bodies of Agaricus bitorquis had a stronger flavour than Agaricus bisporus were consistent with our observations on the 1-octen-3-ol content of these mushroom species (Table 30). Hence we may conclude, that 1-octen-3-ol is a good test substance in examining fungal material for the presence of a mushroom flavour.
5'-GMP was produced by all strains studied, but in varying amounts and there was some difference in the excretion of 5'-GMP into the medium. With several strains (Agaricus bisporus, strain I8; Coprinus comatus, both strains; Lentinus edodes; Morchella esculenta; Pleurotus ostreatus; Volvariella volvacea) most of the 5'-GMP was found in the medium, while the excretion was less with some other strains (Agaricus bisporus, strain A1; Agaricus bitorquis; Beauveria tenella, two strains).
It should be noticed that the production of 5'-GMP by the mushroom species mentioned is much smaller than the production of 5'guanine nucleotides by mutants of Brevibacterium ammoniagenes [50]. This bacterium, which was investigated on the suitability for an industrial production of 5'-guanine nucleotides, formed up to 9.7 g/1 of 5'-GMP, 5'-GDP and 5'-GTP.
The total production of 1-octen-3-ol and 5'-GMP was calculated per 10 g of dry mycelium, grown in submerged culture, and compared with the amounts of 1-octen-3-ol and 5'-GMP per 10 g of dry matter of fresh fruit bodies, which were calculated from the data presented in Table 30, considering the fact that 57 g of dry fruit bodies resulted on an average in one litre of aqueous extract.
With the two Agaricus bisporus strains and with Coprinus comatus (strain L) this relative production (Table 32) of 1-octen-3-ol was much higher in submerged culture than in fruit bodies. Probably the concentrations in the submerged mycelium were too high for a pleasant flavour unless the mycelium was mixed with other food.
aIncluding the amounts released into the culture medium; calculated from Table 31.
bCalculated from Table 30.
cMeasured in dried fruit bodies.
No analytical data for fresh fruit bodies of Lentinus edodes were available for comparison with flavour production in submerged culture. For the other strains mentioned in Table 32, the production of 1-octen3-ol was higher in fruit bodies than in the submerged mycelium.
The production of 5'-GMP by submerged mycelium of Agaricus bisporus and Agaricus bitorquis was practically equal to that in fruit bodies. Submerged Calvatia gigantea mycelium produced much more 5'-GMP than fruit bodies did, whereas submerged mycelium of Coprinus comatus and Pleurotus ostreatus produced less 5'-GMP than fruit bodies did.
5.3. Some factors influencing the production of flavour compounds by submerged mycelium of Agaricus bisporus (isolate A1) and Coprinus comatus (isolate L)
Only little is known about the influence of the culture medium and the incubation time on flavour production in submerged cultures of mushroom mycelium.
Block et al. [16] observed, that mycelium of Agaricus blazei, grown in citrus press water, had a bitter flavour, which could be removed by washing the mycelium thoroughly. Szuecs [139] claimed that addition of lecithine or an edible oil to the culture medium yielded mycelium with a better flavour. Heinemann [68] found that addition of skim milk to the medium improved the flavour of mycelium of some mushrooms.
Some workers [72,139] stated that the mushroom flavour was produced at the end of the growth, during the phase of autolysis. Humfeld and Sugihara [72] claimed that to obtain a mycelium yield with a good strong mushroom flavour it is necessary to continue the fermentation for 1 to 2 days after all the sugar has been consumed and the maximum amount of mycelium has developed. Apart from the fact that these authors worked with Beauveria tenella instead of, as they believed, Agaricus bisporus, their view is scarcely consistent with the fact that young fruit bodies and even primordia have a good flavour. Eddy's experiments [33] did not support the findings of Humfeld and Sugihara. His attempts to age cultures of Coprinus comatus grown in fermentors were unsuccessful since, when all the sugar was used up, the organic nitrogenous compounds in the medium (yeast autolysate) were attacked with the production of an ammoniacal smell.
Szuecs [139] patented a process for the enhancement of the flavour of mushroom mycelium grown under submerged conditions by adding sodium chloride (preferably about 4%) to a washed mycelial product and storing for up to 8 days at 40 °C. Eddy [ 33] tried this process with submerged grown mycelium of Coprinus comatus, Marasmius oreades, Agaricus arvensis and Agaricus bisporus. Only with Coprinus comatus was any increase in flavour noticed, but this was not mushroom like.
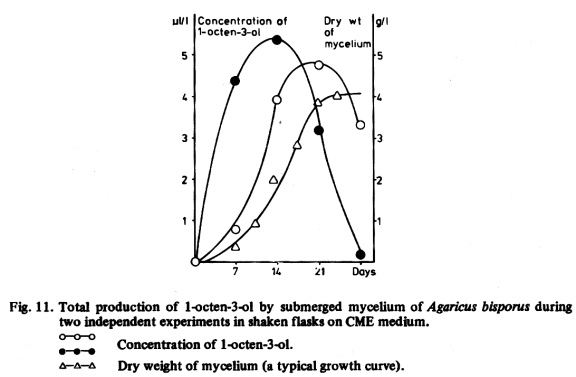
The production of 1-octen-3-ol and nucleotides by our strains of Agaricus bisporus and Coprinus comatus were studied as a function of time. A number of parallel flasks containing the CME medium were inoculated. After different periods of incubation on a shaker, up to 28 days, two flasks were harvested and their entire contents were analysed for 1-octen-3-ol and nucleotides. Fig. 11 shows the production of 1-octen-3-ol by Agaricus bisporus in two experiments of this type. In both tests the concentration of 1-octen-3-ol showed a maximum, but in one the maximum was reached about 7 days later, while the dry weight increased in both experiments in the same way. This shows that the moment at which the production of 1-octen-3-ol starts is not exactly reproducible. The reason for this phenomenon is unknown. With Coprinus comatus similar curves were obtained, but the concentration of 1-octen-3-ol was lower, which agrees with the results presented in Table 31.
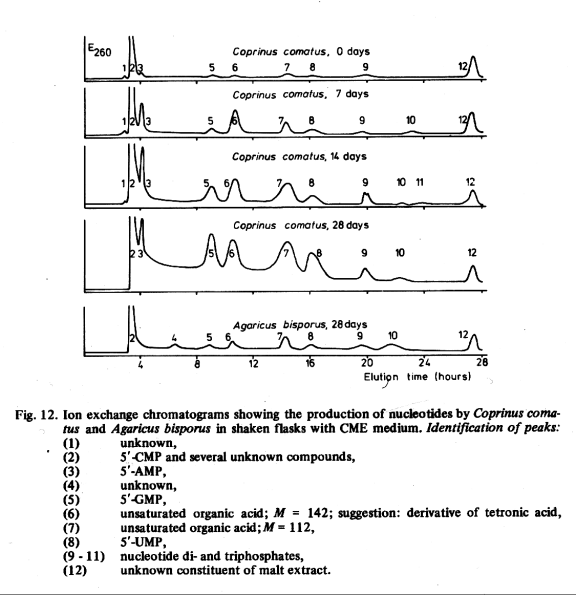
The production of nucleotides appears in Fig. 12. The highest concentrations of 5'-GMP and 5'-UMP in the cultures of Coprinus comatus were observed after 28 days. This might be caused by autolytical breakdown of RNA in the later stages of the growth. The concentration of 5'-AMP showed a maximum after about 21 days. With Agaricus bisporus the highest concentration of 5'-GMP and 5'-UMP' was also observed after 28 days, but the production was much less than with Coprinus comatus. Only the chromatogram of the 28 days old cultures of Agaricus bisporus is presented in Fig. 12. Some peaks (6 and 7) in Fig. 12 could not be identified conclusively. The amounts of substance were small and from the UV, IR and mass spectra it was only concluded that the compounds concerned were unsaturated organic acids. No further investigations of these compounds were undertaken because the purpose of the experiment was only to study the production of nucleotides. Peak 2 was a mixture of 5'-CMP and several other compounds and peak 12 was not identified, since this compound was present in the culture medium.
Comparison of Fig. 11 with Fig. 12 shows, that 1-octen-3-ol attained its maximal production earlier in the growth than 5'-GMP, which reached its maximum at the same time or later than the dry weight of mycelium.
a5 g/l. b30 g/l (Agaricus bisporus) and 60 g/1 (Coprinus comatus).
c2 g/l d3.9 g solids/1 e250 ml/l (Agaricus bisporus) and 500 ml/l (Coprinus comatus).
f2.5 ml/l gMed. = Detected in the medium. hMyc. = Detected in the mycelium.
Table 33 shows the results of some attempts to influence the production of aroma compounds by varying the composition of the culture medium. Malt extract, which is a rather expensive medium, was replaced with glucose plus yeast extract, meat extract or vegetable extract. The influence of Tween 80 was also investigated, because addition of detergents has been shown to enhance the production of guanine nucleotides by Brevibacterium ammoniagenes [49]. It has been suggested that lipids, especially linoleic acid [133] are precursors of 1-octen-3-ol and 1-octen-3-one. For this reason, and in consideration of the statement of Szuecs [139] as regards the effect of an edible oil on the flavour, the influence of olive oil was tested. Because a high flavour intensity has been observed in static cultures [26,33], the production of flavour compounds in such a culture on CME medium was also studied.
The concentrations of 1-octen-3-ol and 2-methyl-2-penten-4-olide were measured after different incubation times up to 23 days. Only the highest concentrations observed in a particular medium are presented in Table 33. These maxima were usually reached at the end of the period, but in some media the maxima were reached earlier. The concentration of 5'-GMP and the dry weight of mycelium were only measured after 14 days.
In none of the media containing a substitute for malt extract the production of 1-octen-3-ol was higher than in the CME medium. For both the production of 1-octen-3-ol and the yield of mycelium, vegetable extract was the best substitute. In all submerged cultures of Agaricus bisporus with a good production of 1-octen-3-ol, this compound was excreted into the culture medium only to a small extent.
Coprinus comatus, when grown in the CME medium, released 1-octen-3-ol into the medium, whereas this compound remained in the mycelium when the strain was grown in CME with addition of olive oil. The production by our strain of 2-methyl-2-penten-4-olide, another important aroma compound of Coprinus comatus, was rather high in most media, and there was a marked excretion into the medium. The greatest total amounts of 2-methyl-2-penten-4-olide were observed in the cultures with vegetable extract. The total amount in these cultures was more than three times as large as in the CME medium.
The production of 5'-GMP by Agaricus bisporus did not vary much with the media, and was significantly lower only in the medium with olive oil in shaken culture. More variation was observed with Coprinus comatus. The highest amount of 5'-GMP was found in the cultures with the vegetable extract, where most of it was released into the medium. In the other media also the mycelium contained 5'-GMP, but the amounts in the media were always greater than in the mycelium.
The static cultures of Agaricus bisporus produced very much 1-octen-3-ol, while the aroma production in the static cultures of Coprinus comatus was rather poor.
Summarising the results presented in Table 33, we may conclude that vegetable extract, which is cheap because it is produced from waste material, can replace malt extract rather well, if a suitable carbohydrate is added. With meat extract and yeast extract, the production of one or more flavour compounds and the yield of mycelium were much smaller. The addition of Tween 80 and olive oil did not cause the expected changes in the maximal production of nucleotides and 1-octen-3-ol, although it stimulated mycelial growth. As was expected, Agaricus bisporus produced much 1-octen-3-ol in static cultures, but most of this flavour component was excreted into the medium.
The experiments described above had to be limited, because of the time-consuming analytical work. No definite conclusions can yet be drawn as regards the influence of the components of the culture medium on flavour production. Little or nothing is known about the biosynthesis of most mushroom flavour compounds; thus no predictions of the influence of the composition of the culture medium on flavour production can be based on fundamental knowledge. For the attainment of such knowledge much microbiological and biochemical research is required.
5.4. The cultivation of Agaricus bisporus and Coprinus comatus in a 10 litre fermentor
For a better comparison of the chemical composition of mushroom mycelium grown in submerged culture with that of fruit bodies, an identification of other volatile constituents of the mycelium than those examined above was considered necessary. The mycelium was therefore grown in a fermentor, so that it was possible to produce an aroma concentrate from about 100 g (dry weight) of mycelium.
The results of some fermentor experiments are shown in Table 34. The compositions of the media were chosen in the expectation that they would provide good yields of mycelium as well as good aroma production. For Agaricus bisporus vegetable extract with addition of glucose, phosphate and casein was used and Tween 80 was supplemented because it stimulated the growth (Table 33). The expected production of nucleotides would be the same as in the CME medium while the expected production of 1-octen-3-ol would be lower, which was not considered to be a disadvantage, because the production of this compound in the CME medium was much higher than in fruit bodies (Table 32). For Coprinus comatus a medium based on skim milk was used. This medium provides good growth, as well as good flavour production [14,148].
aUncertain, since no representative samples of mycelium could be taken from the fermentor before the end of the run, because of the large pellet size.
The agitation of the medium greatly influenced the results of the experiments. With Coprinus comatus the highest yields of mycelium were obtained with a stirring rate of 450 rev/min. Agaricus bisporus grew better at 225 rev/min than at 450 rev/min, giving a higher maximum yield of mycelium while the time required to obtain this yield was shorter. It is interesting, that growth in fermentors was considerably faster than in shaken flasks, where 2 to 3 weeks were necessary to reach the maximum yield of mycelium.
Differences in aeration did not have much influence, suggesting that the rate of oxygen transfer from the air bubbles to the medium was not the limiting factor for the slow growth of the fungi concerned. Van den Berg [14] determined the oxygen uptake by submerged mycelial pellets of Coprinus comatus in a glucose-skim milk-meat extract medium in a fermentor and found values between 3 x 10-5 and 6 x 10-5 mg oxygen per mg dry mycelium per min (mg.mg-l.min-1 ). When the oxygen demand was calculated from the maximal growth rate and Yoxygen (mg oxygen needed for the production of 1 mg biomass) a maximal value of 5.2 x 10-4 mg.mg-l.min-1, which can only be reached during the phase of most rapid growth, was found. Yoshida et al. [163] determined the oxygen consumption by mycelium of Lentinus edodes suspended in 0.9% sodium chloride. Fibrous mycelium used maximally 4.2 x 10-5 mg.mg-1.min-1, while mycelial pellets consumed 1.8 to 3.3 x 10-1 mg.mg-l.min-l. Wikén and Somm [158] determined the oxygen demands for growing young mycelium of Mycelium radicis atrovirens in a glucose-ammonium tartrate medium as 4.8 x 10-4 mg.mg-l.min-1. The endogenous respiration of the washed mycelium in distilled water or phosphate buffer was equal to that shown by the intact mycelium in its actual culture medium. After starvation for 48 h the endogenous respiration was only 2.4 to 7.2 x 10-5 mg.mg-l.min-l. The oxygen consumption of the starved mycelium in 0.056 M glucose was 2.1 to 2.6 x 10-4 mg.mg-1.min-l. No experimental data are available on the oxygen demand of submerged mycelium of Agaricus bisporus, but it seems reasonable that this species needs less oxygen than more rapidly growing species like Coprinus comatus, Lentinus edodes and Mycelium radicis atrovirens.
With the maximal mycelial yield of Coprinus comatus, as found in our present fermentor experiments (9.8 g/1) and the maximum oxygen demand (5.2 x 10-4 mg. mg-l.min-1) the oxygen consumption of the mycelium in the culture broth would be at most 5.1 mg.l-1.min-1.
The rate of oxygen transfer in a fermentor may be expressed in terms of the average driving force for mass transfer [155]
R = KV ( PG,av-PL ),
where R = overall oxygen transfer rate (mg.l-1.min-1),
KV = volumetric oxygen transfer coefficient (mg.l-l.min-l. atm-1 ),
PG,av = average partial pressure of oxygen in gas phase (atm),
PL = partial pressure of oxygen in liquid phase (atm).
The maximum value of R is reached when PL = 0, i.e. when the oxygen consumption of the mycelium is so high that the rate of oxygen transfer limits the growth. This was never observed in our experiments. In the cultures reported in Table 34 the oxygen concentration did not become less than 50% of the saturation concentration. Van den Berg [14] and Van Vliet [148] used a more concentrated medium for the growth of Coprinus comatus, so that the yield was higher. The lowest oxygen concentration they observed was 10% of the saturation concentration.
According to Weinshank and Garver [155] the value of KVV in most industrial fermentors is 1.6 to 8.0 x 102 mg.l-l.min-l.atm-l , while in laboratory fermentors it usually does not exceed 1.6 x 102
mg.l-l.min-l.atm-l ; frequently the value is much lower. In our fermentor experiments, reported in Table 34, KV was between 0.23 and 0.87 x 102 mg.l-l.min-l.atm-l, depending on the rates, of stirring and aeration and on the dry weight of mycelium in the broth. PG,av was at least 0.20 atm.
With the lowest value of KV the maximal rate of oxygen transfer (when PL = 0) was 0.23 x 102 x 0.20 = 4.6 mg.l-1.min-1, which is of the same order of magnitude as the maximum oxygen demand calculated above (5.1 mg.l-l.min-1). This shows that the rate of oxygen transfer probably did not limit the growth. Differences in results apparently caused by differences in aeration are more likely to have arisen from changes in the agitation of the medium brought about by air bubbles than from differences in the oxygen supply.
As was previously found in shake flasks (Table 5), Coprinus comatus also utilised lactose in the presence of glucose in the fermentor, thereby liberating only a small amount of galactose. The efficiency of the sugar utilisation was moderate. When the medium was not stirred too quickly, a little more than two grams of sugar were consumed for the production of one gram of mycelium. The sugar consumption by Agaricus bisporus was perhaps a little more efficient, but it should be borne in mind that in these cultures a small amount of Tween 80, which also serves as a carbon source, was present.
The crude protein content of the mycelium varied from 36 to 57% for Coprinus comatus and from 25 to 32% for Agaricus bisporus. The effect of aeration and agitation on the crude protein content was not clear.
The production of aroma compounds also depended on the rate of stirring. The concentration of 1-octen-3-ol in the Coprinus comatus mycelium markedly decreased below and above the optimum stirring rate (450 rev/min), while the total production of 2-methyl-2-penten-4-olide increased with increasing agitation. A pronounced maximum in the concentration of these two compounds was observed after 4 to 6 days. After 9 days most of the 1-octen-3-ol and 2-methyl-2-penten-4-olide had disappeared. There was no clear correlation between the production of 5'-GMP and the stirring rate. The excretion of 2-methyl-2-penten-4-olide and 5'-GMP into the culture medium was high with fast agitation, possibly because the smaller pellet diameter facilitated their diffusion.
In the fermentor cultures of Agaricus bisporus the production of 1-octen-3-ol, 5'-GMP and mycelium was lower than in shaken flasks (Table 33). We tried to improve these results by using the CME medium (stirring rate 225 rev/min; aeration 1.7 1/min), but no higher production of mycelium or aroma compounds was observed. In our earlier experiments [26] in the Marubishi fermentor we found higher yields (7.8 g/1) in the CME medium supplemented with olive oil.
The formation of pellets by fungi in submerged culture is influenced by various factors, among which the composition, pH and viscosity of the medium, the size of the inoculum and the rates of agitation and aeration are important (see the review of Whitaker and Long [156]). Theoretical models for pellet formation were developed by Yoshida et al. [163] and improved by several other workers. Van den Berg [14] and Van Vliet [148] used a model based on the assumption that the diffusion of oxygen from the medium into the pellet is the limiting factor for growth. When the pellet diameter exceeds a critical value (0.6 mm for Coprinus comatus), the oxygen concentration in the centre of the pellet becomes zero, and the mycelium begins to autolyse. Calculations based on this model were in reasonable agreement with the measured growth of Coprinus comatus in fermentors. Further testing of the model is required however [168].
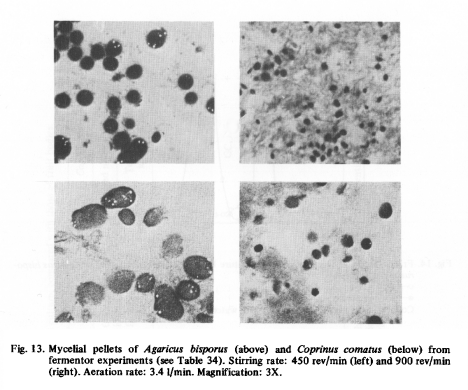
Table 34 shows that increasing the stirring rate from 225 to 900 rev/min at an aeration of 3.4 1/min decreased the average pellet diameter from 2.28 to 0.54 mm. Some pellets are shown in Fig. 13. The pellet size was not distributed normally, but the deviation from normal was rather small. If a normal distribution is assumed, the least significant difference in the average pellet diameter is about 10%. In shaken flasks the average pellet diameter of Coprinus comatus varied from 9 to 0.9 mm when the rotation of the shaker increased from 80 to 300 rev/min [14].
When the pellet diameter was above 1.5 mm it became difficult to take representative samples of mycelium before the end of the fermentor run. With the cultures of Agaricus bisporus it was possible to estimate the growth indirectly by measuring the laccase activity in the medium. Laccase is a phenol oxidase occurring in several higher fungi and is often excreted into the culture medium [98]. Legrand and Martin [91] studied the production of laccase by Agaricus campestris. The enzyme was formed in a complex, but not in a synthetic medium. The synthesis of laccase required copper ions and an unknown factor, which was present in the medium and was also found in coco-nut milk. Molitoris and Esser [104] purified phenol oxidases of Podospora anserina. They isolated tyrosinase (molecular weight 42,000 to 100,000) and three laccases (molecular weights 70,000, 80,000 and 360,000).
In our previous work a correlation was shown to exist between laccase activity and the yield of mycelium in shaken flasks and in the fermentor [26]. Laccase activity depended on the medium and on the rates of agitation and aeration. When these were kept constant, the laccase activity was useful for a rough estimation of the growth in a particular medium. Since laccase rarely occurs extracellularly in moulds, yeasts and bacteria, contamination of the culture by another laccase producing organism is practically improbable. Hence the determination of laccase in the medium is also a qualitative test for the development of Agaricus bisporus, so that it is easy to detect infections in an early stage of growth. Table 34 shows, that laccase activity was maximal at a stirring rate of 450 rev/min and an aeration rate of 1.7 1/min, conditions which are not optimal for mycelial growth.
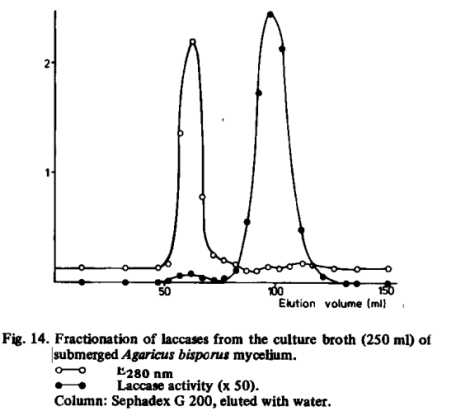
Agaricus bisporus produced at least two extracellular laccases (Fig. 14). The molecular weights were not determined accurately, but from the elution volumes on the Sephadex G 200 column it may be concluded that the first eluting enzyme with only low activity has rather high molecular weight, while that of the other is of the order of magnitude of 105. These enzymes may be the same as or similar to those with molecular weights of 360,000 and 70,000 to 80,000, respectively, which were found by Molitoris and Esser [104].
5.5. Comparison of the flavour compounds in mushroom mycelium grown in submerged culture with those present in fruit bodies
The mycelial pellets of Agaricus bisporus and Coprinus comatus smelled and tasted like mushrooms. During the fermentor experiments described previously Coprinus comatus showed the highest production of both 1-octen-3-ol and mycelium with a stirring rate of 450 rev/min and an aeration of 3.4 1/min (Table 34). The production of 2-methyl-2-penten-4-olide under these conditions was not as high as in the experiments with an agitation of 900 rev/min, although the difference was comparatively small (about 20 to 30%). Consequently the fermentor culture was repeated with a stirring rate of 450 rev/min and an aeration of 3.4 1/min. The mycelium was harvested after 4 days, when the concentration of 1-octen-3-ol and 2-methyl-2-penten-4-olide was maximal. The mycelial pellets thus obtained were used for the preparation of an aroma concentrate.
aGC = Comparison of GC-retention times.
OX = Oxidation with potassium bichromate and oxidised product identified by GC.
RED = Reduction with sodium borohydride and reduced product identified by GC.
MS = identification by mass spectrometry.
baccording to Cronin and Ward [24], Picardi and Issenberg [121], and Wasowicz [154], respectively.
cFrom Table 27
The highest production of 1-octen-3-ol in the mycelium of Agaricus bisporus was found with a stirring rate of 225 rev/min and an aeration of 1.7 1/min. In these conditions the maximum concentration of 1-octen-3-ol in the mycelium was reached after 7 days. The mycelium of this age was used to prepare an aroma concentrate.
The aroma concentrates were analysed; the results are presented in Table 35, where the flavour compounds in the mushroom mycelium from submerged cultures are also compared with those of fruit bodies. There is undoubtedly a great similarity between the volatiles of Agaricus bisporus mycelium and those of fruit bodies. Benzaldehyde, benzyl alcohol, 3-octanol, 3-octanone and 1-octen-3-ol, which have been found in fruit bodies by two or three groups of investigators, were also present in the mycelium. The presence of three more compounds found in fruit bodies by at least two groups (furfural, 2-octen-1-ol and 1-octen-3-one) could not be established in the submerged mycelium but it was not absolutely impossible that 2-octen-l-ol was present; a peak in the gas chromatogram, which might have represented this compound, was too small for a mass spectral identification, while synthetic 2-octen-1-ol was not available as a reference compound for the gas chromatography. 1-Octen-3-one, found in fruit bodies, is possibly an artefact of the isolation procedure (Chapter 4).
There was also a similarity between the volatiles of the submerged mycelium of Coprinus comatus and those of fruit bodies. 2-Methyl-2-penten-4-olide, 3-octanone and 1-octen-3-ol were all present in both materials. In addition there was strong mass spectral evidence of the presence of 2-octen-l-ol in the submerged mycelium. So far we did not find this compound in fruit bodies. 1-Octanol and 3-octanol were not found in the submerged mycelium, while the presence of dodecanol was uncertain. Because 1-octanol and 3-octanol were present in fruit bodies in sufficient amounts to influence the flavour, their absence in the submerged mycelium might have made the flavour of this different from that of the fruit bodies. However, the presence of a substantial amount of 1-octen-3-ol assures that the mycelium has an intense mushroom flavour. 2-Methyl-2-penten-4-olide is a rather unstable compound [4] After its detection in the mycelium as a major volatile, in a concentration that was nearly two times that of 1-octen-3-ol, a much smaller amount was found in the final aroma concentrate.
We may conclude that sensory observations of the flavour of mushroom mycelium are supported by the identification of several flavour compounds. The flavour of the culture medium of Coprinus comatus after growth was stronger than before inoculation. This can be explained by liberation of amino acids (see Table 19) and the production of nucleotides and 2-methyl-2-penten-4-olide (see Table 34), which were released into the medium. However, the flavour of the medium was not mushroom-like and there was an unpleasant, bitter after-taste. The medium must therefore be removed thoroughly before the mycelium is used for flavouring. Most of the 5'-GMP, a great part of which is found in the medium, will be lost in this way. The culture medium of Agaricus bisporus also had a flavour which, although it was not unpleasant, was unlike that of mushrooms; with this organism, too, it is advisable to remove the medium thoroughly from the mycelium. So, although 5'-GMP and glutamic acid are present in the culture medium, their impact on the flavour is diminished because of the presence of off-flavours.
5.6. Conclusions
The investigations described in this chapter have shown that several strains of mushroom mycelium are able to produce flavour compounds in submerged culture, whereas other closely related strains do not produce such substances in noteworthy amounts. The methods used for the 10 litre cultures of Agaricus bisporus and Coprinus comatus were suitable for producing sufficient quantities of mycelium of these fungi for examination of the flavour, but the yields of mycelium were lower than could be expected from the preceding experiments in shaken flasks. The analysis of the volatiles of the submerged mycelium of Agaricus bisporus and Coprinus comatus showed the presence of the most important flavour compounds, but the quantitative relations were not the same as in fruit bodies.
There is no reason to believe that the strains examined in the present work are the best organisms for flavour production. It is quite possible that screening of a very great number of mushroom strains for their ability to form and accumulate flavour compounds might lead to the discovery of organisms with a high production of such compounds, or of organisms which are characterised by a spectrum of aroma substances that is qualitatively and quantitatively more similar to, or even better than that of the corresponding fruit bodies.
REFERENCES TO CHAPTER 5
{kind=link}
{kind=link}
{kind=link}
{kind=link}